EL METABOLISMO
1. GENERALIDADES.
Tanto los organismos unicelulares como pluricelulares, la célula debe hacer
desarrollar sus funciones de relación y reproducción, que se basarán en las
funciones de nutrición, particularmente, en los aportes materiales y
energéticos.
La mayor parte de la dinámica energética celular se establece en torno a un
nucleótido trifosforilado: el ATP o adenosín trifosfato.
2. Anabolismo.
El metabolismo es el conjunto de reacciones químicas que se producen en el
interior de la célula y que conducen a la transformación de los diferentes
compuestos, tanto para proporcionar determinados metabolitos, como para
eliminar sustancias y para cubrir las necesidades energéticas de la célula.
Dentro del conjunto del metabolismo se pueden considerar dos fases:
Destrucción de la materia orgánica -------- CATABOLISMO
Síntesis de la materia orgánica ------------- ANABOLISMO
El anabolismo consiste en la construcción de materia orgánica compleja
a partir de las moléculas sencillas que hay en el citoplasma, requiere energía
en forma de ATP.
Las distintas reacciones químicas del metabolismo se denomina vías
metabólicas y las moléculas que intervienen en ellas se llaman metabolitos.
3. LA FOTOSÍNTESIS
3.1. ASPECTOS GENERALES
Los procesos fotosintéticos son una parte importante del anabolismO en los vegetales, ya que hacen posible la incorporación de sustancias inorgánicas sencillas y su transformación en materia orgánica.
La fotosíntesis, en sentido estricto, es la conversión de energía luminosa en
energía química (ATP) que puede utilizarse para la síntesis de materia orgánica.
Esto es posible gracias a la existencia de moléculas especiales, denominadas
pigmentos fotosintéticos, capaces de captar la energía luminosa.
Se distinguen dos tipos de procesos fotosintéticos:
· Fotosíntesis oxigénica. Es propia de las plantas superiores, las algas y las cianobacterias en las que el dador de electrones es el agua, y en consecuencia, se desprende oxígeno.
· Fotosíntesis anoxigénica. Es propia de las sulfobacterias purpúreas y verdes, en las que el dador de electrones no es el agua, sino, generalmente, el sulfuro de
hidrógeno, por lo que no desprende oxígeno.
Laboratorio virtual: Fotosíntesis Mit.edu
3.2. FOTOSÍNTESIS OXIGÉNICA O VEGETAL
En los organismos que la realizan, los pigmentos fotosintéticos se encuentran en los cloroplastos (en las membranas de los tilacoides), existiendo dos agrupaciones de pigmentos fotosintéticos con proteínas.
Fotosistema I (PSI).
Se localiza en las membranas de los tilacoides, en contacto con el estroma. El centro de reacción contiene dos moléculas de clorofila a denominadas P700 puesto que tienen su punto de máxima absorción a una longitud de onda de 700 nm.
Fotosistema II (PSII). Se localiza en los grana. Su centro de reacción contiene dos moléculas de clorofila a denominadas P680 que tienen su máxima absorción a una longitud de onda de 680 nm.
Estructuralmente, cada fotosistema está a su vez formado por la antena y e centro de reacción.
· Antena: consta de 300 moléculas de pigmentos fotosintéticos, principalmente clorofila a, clorofila b y carotenos, asociados a lípidos y proteínas.
· Centro de reacción: constituido por una molécula de clorofila especial (diana) a la que van a parar los electrones excitados en la antena y que ella transfiere al aceptor primario de electrones.
Según la longitud de onda de la luz se excita un pigmento fotosintético u otro.
Los electrones se transfieren siempre hacia los pigmentos que absorben la mayor longitud de onda, hasta llegar a la molécula diana.
3.2.1. FASE LUMÍNICA DE LA FOTOSÍNTESIS
Tiene lugar en las membranas de los tilacoides, en las cuales se localizan las moléculas de clorofila agrupadas
formando los fotosistemas y el resto de los complejos multiproteicos que participan en la captación de la energía
lumínica.Esta fase depende de la radiación luminosa del Sol.
Los dos fotosistemas son excitados simultáneamente por la luz, pero no están en el mismo nivel de potencial de reducción.
Fotofosforilación no cíclica (con transporte acíclico de electrones)
Cuando incide un fotón (energía lumínica) sobre el fotosistema I, el PS 700 se excita y emite un electrón a una órbita de elevada energía, que es inmediatamente captado por un aceptor que lo cede a la ferredoxina (Fd). Como el potencial de reducción de esta molécula es muy alto, el NADP aceptor los electrones y se reduce a NADPH.
Pero ¿cómo se rellena el hueco dejado por el electrón en el PS 700?
El fotosistema II (PS 680) tiene un máximo de absorción a 680 nm.
La iluminación de este fotosistema provoca la excitación y emisión de electrones, que son captados por una cadena de transporte que los termina por ceder al fotosistema I, rellenando el hueco que se había producido.
Pero es ahora el fotosistema II el que presenta el hueco electrónico.
Este es rellenado por electrones procedentes del agua. Como consecuencia de la ruptura de la molécula de agua (fotolisis del agua), se produce la emisión de dos protones al medio y el desprendimiento de O2.
Esto demuestra que el oxígeno procede del agua.
Se producen así una serie de fenómenos que se representan mediante el esquema Z.
Los fotosistemas I y II actúan conjuntamente y son imprescindibles para que los electrones fluyan del agua al NADP.
Ambos fotosistemas están conectados por medio de una cadena de transporte de electrones, en el que participan la plastoquinona (PQ), el complejo de los citocromo f y la plastocianina (PC).
Se conoce como fotofosforilación a la formación de ATP a partir de ADP + Pi, acoplada al flujo de electrones promovido por la luz.
Del mismo modo que en la membrana mitocondrial se generaba un gradiente de protones, en la membrana del tilacoide se produce también un bombeo de protones promovido por el complejo citocromo f.
Se liberan protones también en la fotólisis del agua y en la formación de NADPH.
El flujo de un par de electrones genera energía suficiente para liberar al estroma entre una y dos moléculas de ATP.
Fotofosforilación cíclica (con transporte cíclico de electrones)
Existe una vía alternativa en el flujo de electrones en la fase lumínica de la fotosíntesis, que no conduce al desprendimiento ni a la formación de NADPH y en la que los electrones son cedidos desde la ferredoxina al complejo citocromo f y no al NADP. A través de la plastocianina, se rellena el hueco del fotosistema I.
Estaría es el flujo de electrones cíclico, para distinguirlo del no cíclico, en el que funcionan los dos fotosistemas.
Las consecuencias de esta alternativa son:
· Funciona sólo el fotosistema I, no el II.
· No se desprende oxígeno.
· No se produce poder reductor NADPH.
· Si se produce una molécula de ATP, gracias a la bomba de protones del citocromo f, es lo que se denomina fotofosforilación cíclica. El flujo electrónico cíclico es una muestra de la versatilidad de la fotosíntesis. Se lleva a cabo en los cloroplastos cuando escasea el NADP y probablemente es una forma de producir ATP para otras funciones celulares. El flujo cíclico es el característico de las bacterias fotosintéticas que no desprenden oxígeno, como las verdes del azufre o rojas.
3.2.2 FASE OSCURA O BIOSINTÉTICA DE LA FOTOSÍNTESIS
El poder reductor (NADPH) y la energía libre en forma de ATP producidos en la fase lumínica de la fotosíntesis se emplearán en la fase oscura para la síntesis de materia orgánica a partir de la reducción del CO2.
El mecanismo de las reacciones de la fase oscura mediante el que las plantas fijan el CO2 atmosférico fue descifrado por Melvin Calvin en 1945, por eso se conoce como ciclo de Calvin.
Las reacciones son catalizadas por enzimas que se encuentran en el estroma del cloroplasto. Al considerar globalmente el ciclo de Calvin:
· Para la fijación de una molécula de CO2 se consumen tres moléculas de ATP y dos de NADPH.
· Si consideramos tres vueltas al ciclo:
· Se utiliza el poder reductor de seis moléculas de NADPH.
· Se utiliza el poder energético de nueve moléculas de ATP.
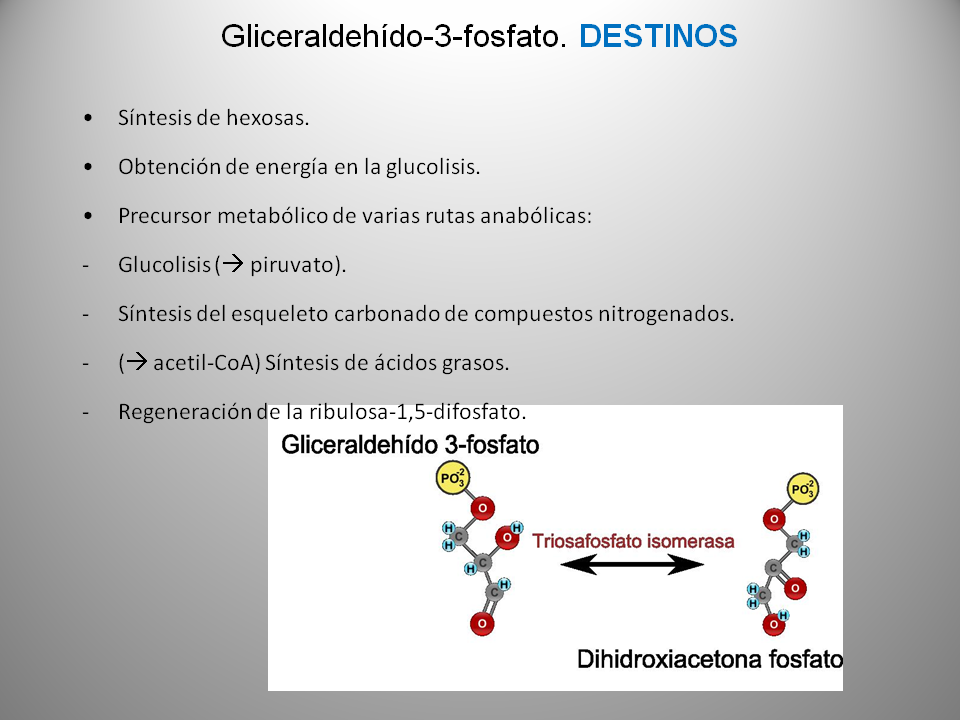
· Se obtiene como producto una molécula de gliceraldehído 3-fosfato (o 3-fosfogliceraldehído).
· A partir de cinco moléculas del gliceraldehído 3 fosfato se regeneran tres moléculas de Ribulosa 1,5 difosfato que, en condiciones apropiadas, se pueden utilizar en un nuevo ciclo.
El resultado final de la fase oscura de la fotosíntesis es la formación de glucosa a partir de CO2.
Las hexosas formadas (glucosa) son precursores de la sacarosa y del almidón productos finales de la fotosíntesis.
Las plantas pueden tener un problema para realizar la fase oscura de la fotosíntesis debido a una imperfección catalítica de la ribulosa 1,5 difosfato carboxilasa (rubisco).
La enzima que fija el CO2 a la ribulosa1,5-difosfato es, además de una carboxilasa, una oxigenasa, lo que significa que puede oxidar también al sustrato, utilizando para ello el mismo centro activo que para su acción como carboxilasa.
El hecho de que actúe con una u otra acción catalítica depende de las concentraciones relativas de O2 y de CO2 en el estroma del cloroplasto en un determinado momento.
En las hojas de las plantas,el intercambio de gases se realiza mediante unos orificios llamados estomas que están relacionados con células especializadas
que permiten el control de apertura y cierre, lo cual depende de algunos factores, entre ellos, la cantidad de agua.
Cuando el agua escasea en condiciones de sequedad y de altas temperaturas, el estoma se cierra, evitándose así la perdida de agua.
Pero también hace que no entre CO2 del exterior y que el O2 producido por fotosíntesis se acumule en el interior.
En estas condiciones, la Ribulosa 1,5 difosfato carboxilasa oxidasa pone en acción su actividad oxidasa y se inicia un proceso dependiente de la luz que, por consumir O2 y producir CO2 se llama fotorrespiración.
La fotorrespiración es, un proceso sin rendimiento ni ganancia en el que un carbono orgánico se convierte en CO2 sin producirse NADPH, ni ATP u otra molécula rica en energía: es, por tanto, perjudicial para la planta, al reducir la eficacia de la fotosíntesis.
Algunas plantas solucionan el problema de la fotorrespiración con una ruta alternativa de fijación del CO2: la vía del C4, llamada así porque las principales moléculas intermediarias son de cuatro carbonos.
También se llama ruta de Hatch y Slack, por sus descubridores.
4. FACTORES QUE INFLUYEN EN LA FOTOSÍNTESIS
· INTENSIDAD LUMINOSA· EL COLOR DE LA LUZ.
· LA CONCENTRACIÓN DE OXÍGENO.
· LA CONCENTRACIÓN DE DIÓXIDO DE CARBONO.
· TEMPERATURA.
· DISPONIBILIDAD DE AGUA.
5. LA QUIMIOSÍNTESIS
5.1. ASPECTOS GENERALESLos organismos capaces de sintetizar materia orgánica a partir de compuestos inorgánicos son autótrofos.
Los fotosintéticos utilizan la luz como fuente de energía necesaria para llevar a cabo este proceso.
Otros, los quimiolitotrofos, realizan Quimiosíntesis.
La quimiosíntesis consiste en la obtención de materia orgánica a partir de inorgánica y de la energía desprendida de reacciones químicas redox exergónicas (que liberan energía).
Este proceso está restringido a un reducido número de especies bacterianas.
5.2. FASES DE LA QUIMOSÍNTESIS
· Obtención de energía. Se obtiene de las reacciones inorgánicas en las que se produce una oxidación que
desprende energía en forma de ATP y NADH.
· Producción de materia orgánica. El ATP y NADH obtenidos en la fase anterior se utilizan para la síntesis de la
materia orgánica por medio del ciclo de CalvinLOS ORGANISMOS QUIMIOLITÓTROFOS
Los quimiolitótrofos presentan una serie de características que definen su actividad quimiosintética.
· Son solo procariotas.
· Viven de una fuente estrictamente inorgánica.
· La fuente de energía es una reacción específica.
· Son aerobios.
· Sintetizan la materia orgánica por medio del ciclo de Calvin.
Este tipo de bacterias se agrupan e identifican por el tipo de oxidación de la que extraen la energía:
· Bacterias nitrificantes: utilizan como sustrato compuestos de nitrógeno. Se distinguen: Nitrosomonas, Nitrobacter.
· Sulfobacterias: oxidan azufre o compuestos reducidos de azufre.
· Ferrobacterias: son abundantes en las charcas de agua dulce con sales reducidas de hierro
































































Cuestiones sobre metabolismo
Entra en joinmyquiz.com e introduce este
código 53549716 .
Enlace URL: https://quizizz.com/join?gc= 53549716
Cuestiones sobre metabolismo
Entra en joinmyquiz.com e introduce este
código 53549716 .
Enlace URL: https://quizizz.com/join?gc=53549716
No hay comentarios:
Publicar un comentario